
A seleção natural é o fenômeno biológico que favorece a sobrevivência de parte da população, ou seja, aqueles seres que, graças à variabilidade genética, herdaram combinações gênicas mais adaptativas a uma determinada condição ecológica. Lembramos que a expressão “mais adaptada” se refere à maior probabilidade que um determinado organismo apresenta de sobreviver e deixar descendentes em certo ambiente. Ao contrário da mutação, a seleção natural atua reduzindo a variabilidade gênica. Assim sendo, em face de haver seleção de apenas alguns genótipos, quanto mais intensa for a seleção natural, menor será a variabilidade gênica na população.
Para se ter uma ideia da atuação da seleção natural, se os tentilhões do arquipélago de Galápagos encontrassem somente sementes grandes e duras, provavelmente apenas aqueles com bicos maiores e mais fortes seriam capazes de sobreviver e, geração após geração, se tornariam mais comuns na população. A seleção de uma característica, seja o tamanho do bico ou qualquer outra, depende da transmissão de genes daquela característica para os descendentes.
Em face de o ambiente não representar um sistema constante e estável, ocorrem diferentes interações entre os organismos e o meio. A heterogeneidade temporal e espacial propiciam diferentes pressões seletivas sobre o conjunto gênico da população, evitando, por vezes, a eliminação de determinados genes que, em condições ideais, não seriam mantidos. É o que acontece com a manutenção na população humana de certos genes que normalmente seriam eliminados por serem pouco adaptativos. Um exemplo é o gene que causa a anemia falciforme (anemia drepanocítica ou siclemia), que estudaremos mais adiante em “seleção estabilizadora” e cuja frequência é alta em extensas regiões da África, onde se verifica uma grande incidência de malária.
TIPOS DE SELEÇÃO NATURAL
Tendo por base os efeitos que a seleção natural exerce nas populações, ela pode ser classificada em três tipos básicos: direcional, estabilizadora (ou normalizadora) e disruptiva (ou diversificadora). O gráfico abaixo mostra como se distribui, geralmente, as várias classes fenotípicas de uma população padrão. Nesse caso, os fenótipos “medianos” são os de maior frequência. Procure relacioná-lo com os gráficos associados aos três tipos de seleção (direcional, estabilizadora e disruptiva), que estudaremos a seguir.

1. Seleção direcional
Nesta seleção, as condições ambientais favorecem um fenótipo extremo, diferente do que representa a média da população (gráfico a seguir), sendo por isso que a frequência alélica muda continuamente na mesma direção. Ela ocorre, portanto, quando a população está sujeita à alterações ambientais em um dado sentido, produzindo, como consequência, mudança regular da população em uma mesma direção. A resistência de insetos a inseticidas, a resistência de bactérias a antibióticos e o melanismo industrial são exemplos de seleção direcional.

Ia – Resistência de insetos a inseticida
Numa população de insetos nitidamente sensíveis à ação de determinado inseticida (DDT, por exemplo), há alguns indivíduos naturalmente resistentes. Essa capacidade de resistir ao agente químico constitui uma característica determinada por certos genes mutantes, que conferem aos insetos que são seus portadores uma variação “favorável”, no caso de a população ser exposta ao inseticida. Dessa forma, pulverizando-se DDT (figura abaixo) sobre esses insetos, percebe-se, inicialmente, uma grande diminuição na população, devido à eliminação dos indivíduos sensíveis. Os insetos resistentes, entretanto, não apenas sobrevivem como continuam a se reproduzir, gerando descendentes dotados do mesmo poder de resistência. Dessa forma, após algumas gerações, a população é constituída, praticamente, apenas de insetos resistentes, levando a que o número de indivíduos permaneça mais ou menos constante, mesmo quando novas aplicações de inseticida são feitas. A população está, portanto, adaptada à presença do inseticida, pois a seleção fixou a variação “favorável”, que confere a capacidade de resistir ao agente químico. Com base no exposto é incorreto dizer os insetos adquiriram resistência ao inseticida, já que se constitui um conceito Lamarckista.
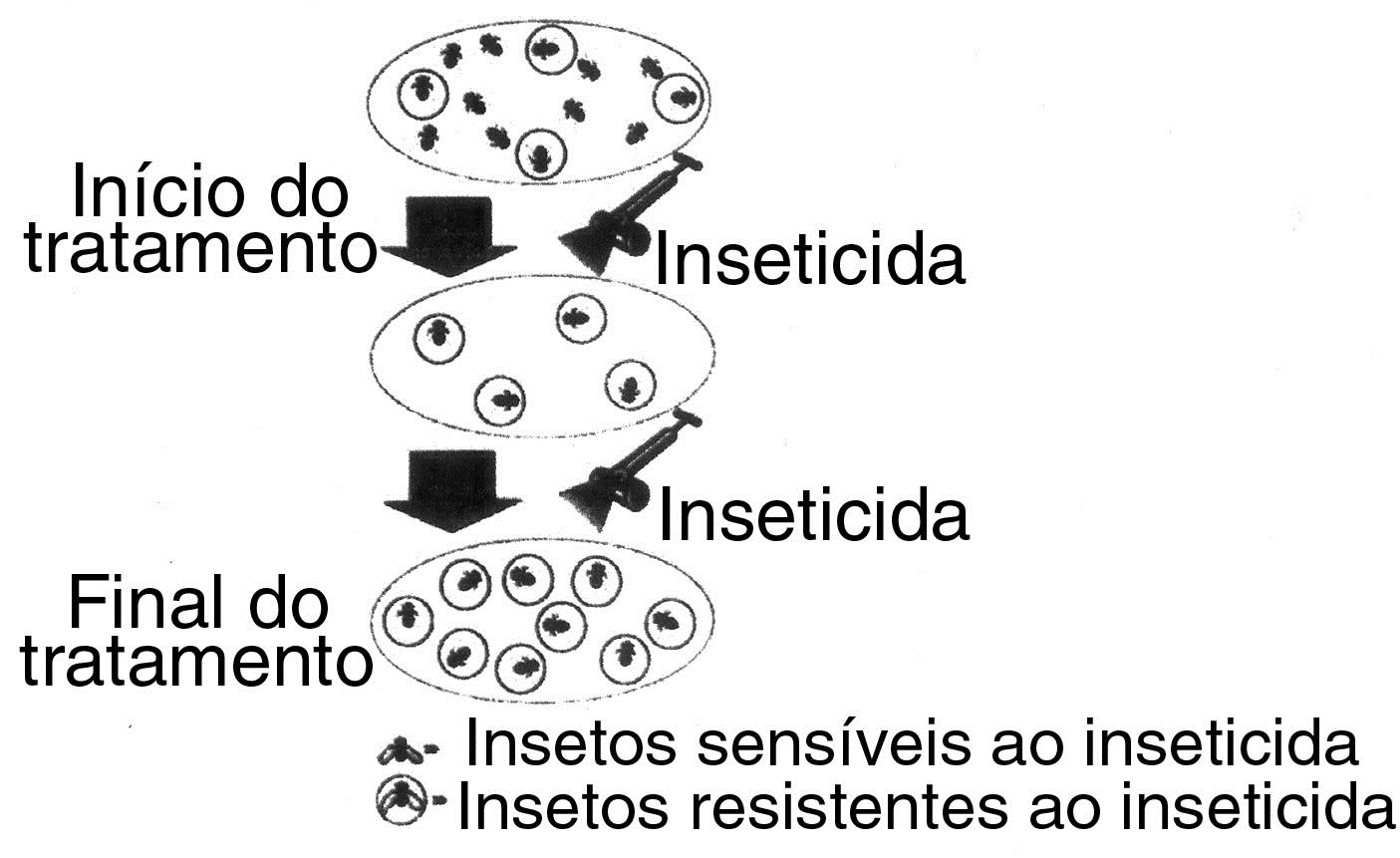
Lembramos que o DDT (sigla de Dicloro-Difenil-Tricloroetano) começou a ser utilizado, em grande escala, na Segunda Guerra Mundial, visando combater os piolhos que transmitiam o tifo e a malária, responsáveis pela morte de milhões de pessoas. Embora o DDT tenha sido sintetizado em 1874, suas propriedades inseticidas, contra vários tipos de artrópodes, só foram descobertas em 1939 pelo químico suíço Paul Hermann Müller, que, por essa descoberta, recebeu o Prêmio Nobel de Medicina de 1948. Em função dos bons resultados obtidos, ele passou a ser empregado, também, na agricultura, com o intuito de combater as pragas agrícolas. O DDT, entretanto, se mostrou nocivo à saúde humana e ao equilíbrio ecológico.
1b. Resistência de bactérias a antibióticos
A exemplo dos inseticidas, os antibióticos podem perder sua eficácia no combate a determinados tipos de bactérias. Habitualmente se diz que as bactérias adquirem resistência ao antibiótico, o que não é correto. Aqui cabe o mesmo raciocínio usado para explicar a resistência dos insetos ao DDT. O que realmente ocorre é a seleção de linhagens bacterianas geneticamente resistentes ao antibiótico que, sobrevivendo, se multiplicam e passam a predominar no grupo. Os antibióticos, da mesma forma que o DDT, funcionam, portanto, como agentes seletivos e não como agentes mutagênicos. Sua ação consiste, em última análise, em eliminar os organismos portadores de genótipos sensíveis, favorecendo os dotados de genótipos resistentes, que surgem por mutações ao acaso (figura a seguir). Neste contexto, já foram selecionados um grande número de linhagens bacterianas dotadas de resistência a vários antibióticos. Reiteramos que essa resistência se deve a mutações que as populações bacterianas sempre estão sujeitas. O mesmo pode ser dito em relação à resistência dos insetos aos inseticidas, que mencionamos acima, e ao melanismo industrial que abordaremos a seguir.

1c. Melanismo industrial
Outro exemplo de seleção natural direcional ocorreu com as variedades clara e escura de mariposas da espécie Biston betularia, cujas populações sofreram, nos últimos 150 anos, mudanças evolutivas profundas com relação à cor. Em várias regiões inglesas, antes da industrialização, rochas e troncos de árvores eram cobertos por liquens, e os bosques próximos às cidades eram ambientes claros. Nessa condição, as mariposas claras existiam em grande número, pois elas se confundiam com os troncos claros das árvores (figura A, a seguir), sendo menos visíveis que as escuras, que se destacavam e eram facilmente capturadas pelos pássaros predadores, sendo, portanto, raras. Com o incremento da industrialização, a fumaça e a fuligem, produzidas pelas indústrias, enegreceram, lentamente, os troncos das árvores e eliminaram os liquens. A partir de então, a variedade escura, também conhecida como melânica, e observada pela primeira vez por volta de 1850 nas proximidades do distrito de Manchester (Inglaterra), teve na sua cor uma proteção contra os predadores (figura B, abaixo), tornando-se cada vez mais numerosa. A variedade clara, por outro lado, tornou-se alvo fácil para a ação predatória dos pássaros, tornando-se cada vez menos numerosa.

Como se pode constatar nesse exemplo, a mudança gradativa de uma condição ambiental determinou alteração na frequência gênica de uma população em um determinado sentido, como resposta à seleção natural. Na atualidade, esse fenômeno se encontra generalizado e é conhecido como melanismo industrial.
Lembramos que a seleção artificial, feita pelo homem com o objetivo de promover o melhoramento genético de espécies animais e vegetais em seu próprio benefício, é também um exemplo de seleção direcional. O mesmo raciocínio se aplica à clonagem, que vem sendo feita pelo homem com o intuito de preservar algumas características desejáveis.
2. Seleção estabilizadora ou normalizadora
Esta seleção atua em populações que vivem em ambientes relativamente estáveis, nas quais a média dos indivíduos está bem adaptada às condições ambientais. Ela favorece indivíduos portadores de formas intermediárias e elimina os que apresentam formas extremas da curva de distribuição normal (gráfico a seguir). A seleção estabilizadora mantém, portanto, a população geneticamente constante.

O favorecimento dos heterozigotos para o gene da anemia falciforme (anemia drepanocítica ou siclemia), em regiões onde há grande incidência de malária, é um bom exemplo de seleção estabilizadora. Esse gene mutante leva à formação de moléculas anormais de hemoglobina e consequente malformação das hemácias, que assumem forma semelhante à foice (figura abaixo), com reduzida capacidade de transportar oxigênio. Dessa forma, em condições ambientais normais, o gene para a siclemia sofre uma forte seleção negativa, levando a que sua frequência seja baixa nas populações em geral.Lembramos que a siclemia é uma anomalia genética de herança autossômica, determinada por um gene letal em dose dupla, comum entre os africanos e pertencente a um grupo de doenças chamadas hemoglobinopatias. Para maiores detalhes acerca da mutação que leva à formação das hemácias falciformes ver “mutação gênica”, matéria publicada neste blog no dia 15/04/2011.

Os indivíduos homozigóticos para a siclemia (ss), apresentam elevada anemia e, via de regra, morrem antes da fase reprodutiva. A tendência, portanto, é que o gene s desapareça, praticamente, da população. Em locais onde a malária é endêmica, entretanto, a frequência desse alelo (gene s) é grande, bem acima do esperado. Essa elevada frequência se deve ao fato das pessoas heterozigóticas (Ss), conquanto sejam ligeiramente anêmicos, pois produzem tanto hemoglobinas normais quanto falciformes, apresentarem maior resistência à malária que os indivíduos homozigóticos normais (SS). Sabe-se que, quando o Plasmodium (protozoário que provoca a malária) penetra nas hemácias consome oxigênio. Com a diminuição de oxigênio, as hemácias siclêmicas se deformam e são fagocitadas pelos leucócitos, antes que o protozoário presente no seu interior se reproduza e se espalhe pelo organismo.
Em função, portanto, dos homozigóticos para siclemia (ss) tenderem a morrer de anemia e os homozigóticos normais (SS) terem elevada tendência a morrer de malária, como vimos acima, os heterozigóticos (Ss) apresentam maior chance de sobrevier e de se reproduzir, transmitindo o alelo mutante s para a geração seguinte. É, em última análise, essa superioridade adaptatiiva dos heterozigóticos que propicia a alta frequência do gene s na população malarígena. Do exposto, é de se supor que a erradicação da malária eliminará a desvantagem dos homozigóticos SS em relação aos heterozigóticos Ss, eliminando, progressivamente, o alelo s, já que os homozigóticos ss continuaram a morrer de anemia.
Outro exemplo de seleção estabilizadora foi verificado em uma população de pardais, a partir da análise do tamanho das asas dos indivíduos mortos após uma forte tempestade. Entre os mortos, a grande maioria era de pardais com asas muito longas ou muito curtas em relação à média da população. Como se pode constatar, a tempestade atua como fator seletivo, eliminando, mais intensamente, os pardais dotados de asas de tamanhos extremos, preservando os que possuem asas de tamanho intermediário. A pressão seletiva favoreceu, em última análise, a sobrevivência dos tipos médios.
3. Seleção disruptiva ou diversificadora
Esta seleção ocorre quando uma população, já adaptada a um ambiente homogêneo, é submetida a diferentes pressões seletivas, de modo que são selecionadas formas adaptativas em direções opostas. Ela favorece, portanto, os indivíduos portadores de formas extremas de certas características, em detrimento das formas intermediárias, gerando dois grupos dominantes na população (gráfico abaixo). Quando esse processo opera, os indivíduos dos extremos da distribuição tendem a produzir maior descendência do que aqueles do centro da distribuição. Admite-se que em face da capacidade que essa seleção apresenta de promover a diversificação de uma população, ela represente a primeira etapa para a formação de novas espécies, processo denominado especiação.
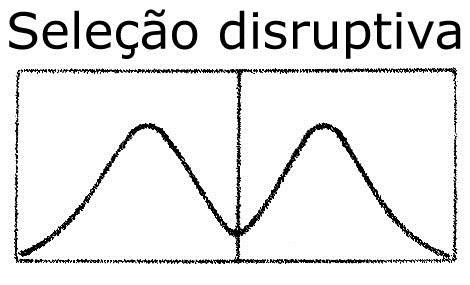
Como exemplo de seleção disruptiva, citamos os besouros que se alimentam de sementes e desprezam as de tamanho pequeno e grande, preferindo as de tamanho médio. Em função disso, serão favorecidos organismos dotados dos fenótipos extremos e, portanto, de plantas produtoras de sementes pequenas e de sementes grandes.
Outro exemplo diz respeito ao crescimento de plantas em regiões próximas de minas de zinco e chumbo. Nas áreas onde os rejeitos são lançados há um elevado nível de contaminação, em contraste com outras áreas não contaminadas. Uma população de plantas mutantes que conseguem sobreviver no solo contaminado leva nítida desvantagem ao crescerem em solo não contaminado, em face da grande competição com a população original de plantas não mutantes lá estabelecidas. Por outro lado, as plantas originais estão restritas às áreas não contaminadas e somente nelas sobrevivem. A diversificação possibilitou ampliar as áreas de crescimento com uma seleção disruptiva que levou o desenvolvimento de dois tipos distintos de plantas em diferentes aspectos. Um deles adaptado a solos contaminados e outro adaptado a áreas não contaminadas.












0 comments:
Postar um comentário